





Evolution and Extinction of Mammals on Islands
Evolution of Island Mammals: Adaptation and Extinction of Placental Mammals on Islands, 2nd Edition. Wiley-Blackwell (Oxford, UK), 2021 (ISBN 9781119675730, eISBN 9781119675716, 576 pp., index, 153 figs, 36 full-colour plates)
More information at Wiley-Blackwell
Evolution on islands differs essentially from evolution on mainlands. Especially islands of the past are uniquely intriguing. Due to millions of years of isolation, exceptional and sometimes bizarre mammals evolved, such as pig-sized elephants and hippos, giant rats and gorilla-sized lemurs, formidable to their mainland ancestors.
Evolution of Island Mammals, Second Edition, provides an updated and expanded overview of the current knowledge on fossil island mammals worldwide, ranging from the Oligocene to the onset of the Holocene, with examples of the fragmentary Cretaceous record. The book addresses evolutionary processes and key aspects of insular mammal biology, exemplified by a variety of fossil species.
Readers familiar with the first edition will find here a host of updated and enhanced material, including:
An entirely new chapter on the island rule
Updated and expanded theoretical chapters
Updated and improved taxonomic information
Extensive coverage of new discoveries
Body masses or body size indices for most extinct island mammals
New figures visualizing the richness of the fossil record
This accessible and richly illustrated textbook is written for graduate level students and professional researchers in evolutionary biology, palaeontology, biogeography, zoology, and ecology.
This material is based upon work published by Wiley-Blackwell, 2021. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the authors and do not necessarily reflect the views of the publisher.

Dispersal and Isolation
The composition and nature of island faunas depend on many factors. These are, amongst others, the isolation and size of the island, its physiography and latitude, the dispersal abilities of its colonisers, the faunal composition as a whole and that of the nearest mainland and the nature of the colonising species. Naturally, these factors are interwoven and subject to changes over time, influencing each other constantly. For example, the physical geography of the island together with local and global climate put a heavy stamp on island size in terms of suitable surface area. A new arrival may have a dramatic impact on resident species.
How to reach an island? Different taxa may disperse differently, so not all dispersal types apply to all taxa.
Three main types of dispersal routes can be distinguished, being over land, over water and through the air. The first type has no restriction, except for strictly aquatic taxa. The second type is open only for taxa that can swim, float or raft on a floating mass across wide water barriers, while the last type is restricted to bats. For invertebrate taxa, hitchhiking on a host during the journey has been recorded, for example freshwater snails sticking to the feathers of a bird, but this kind of dispersal is unknown for mammals. The total distance may be broken up into smaller units, for which the popular term island hopping is sometimes used.
Dispersal over land can be either a corridor dispersal or a filter dispersal. For corridor dispersal, faunal interchange between two areas is possible for all species. For filter dispersal, this is probable for some species but improbable for others. Corridor dispersal takes place over land (land bridge), whereas filter dispersal may include minimal distances over water (step stones, not to be confused with island hopping) or marshy areas.
Continental shelf islands are typically colonised by a normal mainland fauna through corridors (at the start) or filters (in a later stage). During further disconnection and isolation, the filter becomes stronger and eventually no new direct influx takes place anymore. At this point the taxa are confined to the island and may undergo so-called vicariance, the effect of being separated from the rest of the original group by a geographic barrier. This separation often results in a differentiation into new varieties or species. In case an array of new species arises, the term adaptive radiation is applied. Examples of such vicariance are the marsupials of Australia, the fauna of Middle Pleistocene Java and perhaps the ground sloths of the West Indies.
The opposite, continentalisation, happens when an island gradually becomes connected to the mainland. A mainland fauna arrives, first through filter dispersal, later through corridor dispersal. An example is Japan, which was invaded during the Late Pleistocene by a mainland fauna with giant Chinese deer (Sinomegaceros), resulting in the extinction of the native fauna with dwarf elephants (Elephas naumanni).
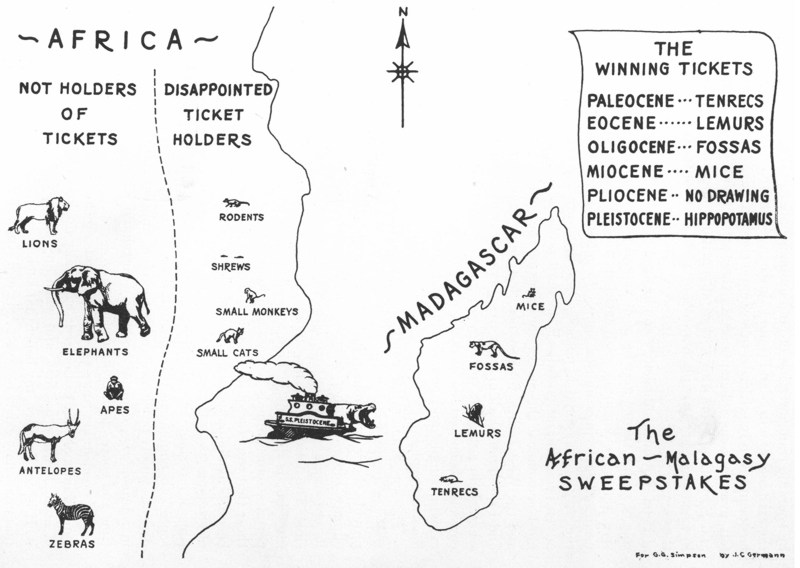
Oceanic islands can only be reached over water or through the air. This type of islands was never connected to the mainland, as is the case with continental shelf islands. This high degree of isolation and the limited dispersal abilities of most species results in a low number of founding species. Freed from the pressure of competition and predation, these founders often evolve in new species, or arrays of new species (adaptive radiation), formidable to their mainland ancestors. The most extreme examples of evolution in isolation are found on these islands. They include pony-sized elephants, giant shrews, astonishing radiations of murids (mice and rats) and diminutive hominids.
Biodiversity
On most oceanic islands, the taxonomic diversity is poor, with many major groups absent (impoverished, depauperate and/or unbalanced faunas). At the same time, many insular founder species have undergone adaptive radiations to fill in the gaps. The lack of diversity on higher taxonomic levels is balanced as it were by high diversity on lower taxonomic levels, as compared to the mainland. Another typical feature of an impoverished fauna are niche shifts. For exmaple, on islands where herbivorous mammals are lacking, large birds take over this ecological niche: the moa on New Zealand and the geese of Ibiza.
Typical insular faunas are devoid of terrestrial mammalian carnivores (unbalanced faunas). This lack of mammalian carnivores leaves the niche of top predator open, which is subsequently filled by birds of prey, such as the giant eagles Garganoaetus freudenthali on Gargano and Haast eagle (Harpagornis moorei) in New Zealand, or reptiles, such as the Komodo dragon (Varanus komodensis) on Flores, Rinca and Komodo and crocodiles on Luzon, Cuba and many other islands.
There is a difference in the understanding of the term balanced between palaeontology and ecology. The way the term is used by most palaeontologists means that large carnivores are present and that thus all major trophic niches are occupied. In ecology, a balanced fauna means it is ecologically stable, for instance the trophic niches are not only occupied, but occupied in ratios which do not encourage transitions.

The Human Impact
The timing and cause of (almost) complete extinction of endemic fossil faunas, termed faunal turnovers, or of a single endemic taxon, termed species extinction, is often poorly known, due to the fragmentary nature of the fossil record and lack of absolute geochronological datings. The subject is, however, of great importance, mainly because of the extensive and ongoing Holocene extinctions of islanders often related to human activity, directly as well as indirectly, as well as climatic changes. Knowing the past, may help us protect the future.
There are two main causal pathways to extinctions of island endemics. Firstly, the natural dynamics of the island system: These include competition with new species (new arrivals or locally evolved species), natural habitat changes (such as related to climate or tectonics), natural disasters and the end of the island (by total submersion or the connection to the mainland). Evidence of these factors is extremely limited in the fossil record. Secondly, the impact of humans. This can be manifold (hunting, active and passive introductions of alien taxa, spread of pathogens, habitat destruction etc.) and acts on levels from the individual taxon to entire faunas. The colonisation of islands by Palaeolithic and Neolithic humans is relatively well documented by archaeological findings (e.g. Flores, Ryukyu Islands, Cyprus, Caribbean islands). Their impact seems limited, and no evidence could be found for widespread island extinctions after Pleistocene hominin arrival. The impact of colonisations during historical times is known in more detail. This impact was often severe, followed by extinctions worldwide.
Extinction events are often caused by a combination of factors, and rarely by a single factor alone. In addition, natural factors and anthropogenic factors are often difficult to separate, for example, whether a competitive species came on its own to the island or was introduced by humans, the eventual outcome is the same. Also, the combination of natural factors and anthropogenic factors may give the fatal blow to a species, for example climate change and hunting.
The dramatic and devastating impact of hunting by first settlers in historical time is generally referred to as Blitzkrieg extinction: fast, overwhelming and with no change for defence. The theory of Blitzkrieg extinction has been used to explain the extinction of the megafauna of New Zealand, Australia, Madagascar and the Americas practically immediately after the arrival of humans. The basic assumption here is that the endemic fauna is naive by nature and thus easy to hunt down. The theory does not include the effects of climate change. The climate of Australia and Madagascar became hotter and drier and large parts lost their original vegetation cover. The model likely explains the sudden extinction events such as that of the moas of New Zealand within a century after the arrival of the Polynesian settlers, but not all.
For example, extinctions in the West Indies started after the arrival of the first Amerindians, some 6000 years ago. However, the Amerindians had no devastating impact on the entire fauna and many taxa continued to exist after their arrival. Thus, the Blitzkrieg scenario is not supported by facts here. On the contrary, the extinction caused by the Amerindians and later by the Europeans presumably took place within a rather broad time frame, sometimes referred to as Sitzkrieg, as opposed to Blitzkrieg. Extinction of West Indian mammalian taxa was likely caused by the culminating effects of centuries of human-induced distress.

Speciation Through Time
Evolutionary changes in island mammals are predictable to a certain extent. In the theoretical chapters of the book, observed anatomical and morphological patterns or trends in island mammals are discussed, and the mechanisms explained.
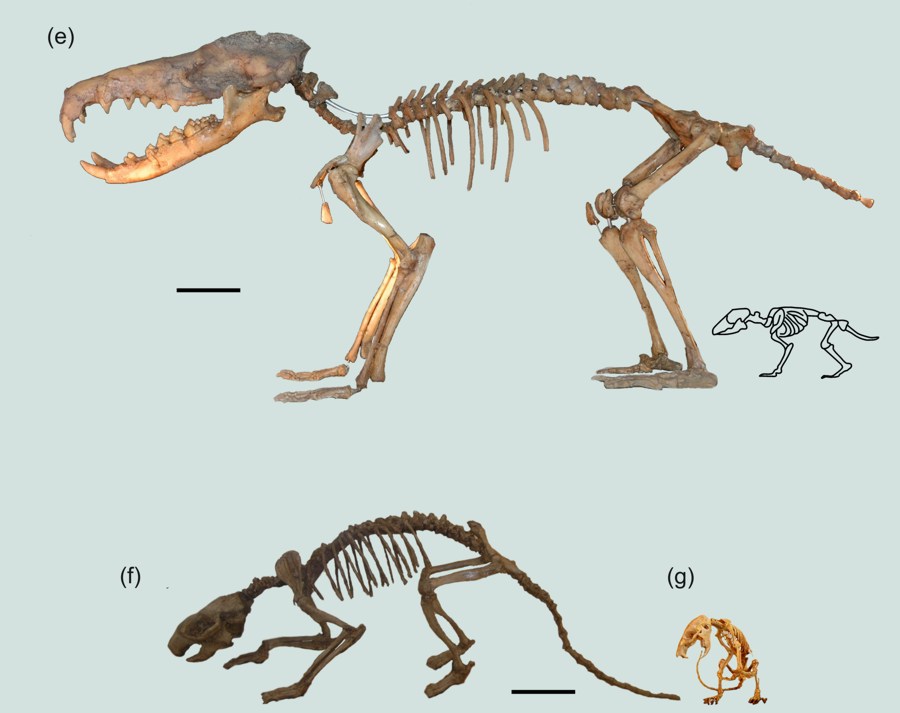
General trends are a change in body size (size reduction in larger mammals, size increase in small mammals), referred to as the island rule, and an increase in hypsodonty in herbivores. Another general trend is the development of a stable, sure-footed but slow type of locomotion (short limbs, bone fusions) in even-toed mammals and proboscideans. Much less known trends (because they are observed in fewer taxa) are the development of ever-growing rodent-like incisors in some goat-like species, the loss of teeth, the simplification of antlers, and more stereoscopic view.
Terms like sympatric and allopatric speciation, metapopulations, species flock and adaptive radiation are explained in the context of island endemics and discussed with examples. Island arches and archipelagos are described, and the speed of evolution of islanders is illustrated with examples like that of a long-term canid lineage (Cynotherium), spanning the entire Pleistocene of Sardinia-Corsica.
Which Animals?
The majority of fossil insular mammals are large herbivores (deer, bovids, hippos, proboscideans, sloths) and rodents. Less frequently found are otters, pigs, hares, rabbits, and insect-eaters. Rare insular mammals are carnivores (canids, hyenas, felids), rhinos and primates. The rarest are bats, but this is due to the fragmentary nature of their fossil record.
This is applies of course only to a global scale. Per island the picture may differ essentially. For example, primates are not rare at all on Madagascar, where they form half the macro-biodiversity. In the remote past, there were even more of them, including gorilla-sized lemurs, but they went extinct some time after humans arrived. On Ibiza, there is a geological time-slice with only bats and birds. For some unknown reason, most likely strong currents, the island could not be reached but by air. Sloths and caviomorph rodents are common in the West Indies, but totally lacking elsewhere. Endemic hamsters are found only in the Mediterranean (Gargano, Mallorca).
Fossil islanders can be roughly divided into two classes: those that arrrived either by swimming or by rafting (sweepstake dispersal or 'by chance' dispersal), and those that became isolated after the region got separated from the mainland (vicariance). Generally, deer (e.g. Candiacervus of Crete, see photograph), hippos, proboscideans, murids belong to the first class; bovids, lagomorphs, primates to the second class. The book describes and discusses the effects of insularity on these taxa, based on what is known from over 300 endemic insular taxa, which lived or still live on at least 80 islands worldwide, from the Late Cretaceous to the Holocene.
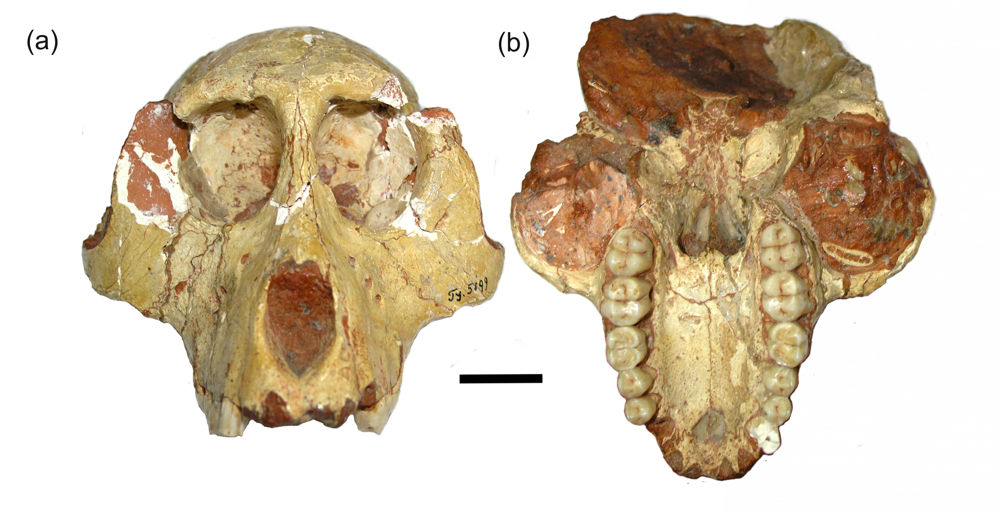
Endemic faunas generally comprise only large or medium-sized herbivores (deer, bovids, proboscideans, hippopotamuses), a range of small mammals (e.g. rats and mice, insectivores, tenrecs, hutias, hares and rabbits), a very limited number of carnivores (mustelids, and incientally, dogs, grey foxes, cats) and primates. Island hominids include the dwarf form of Flores Homo floresiensis.
The discussed islands and island species are found all over the world, and include those of the Mediterranean Sea (e.g. Crete, Sicily-Malta, Cyprus, Dodecanesos, Cycladic Islands), insular Southeast Asia (e.g. Java, Flores, Timor, Philippines, Japan, Ryukyu), the Pacific Ocean (Galapagos, Channel Islands), the Atlantic Ocean (Falkland Islands, West Indies, Canaries), and the Indian Ocean (Madagascar).
Acknowledgments
Our book is dedicated to the memory of the late Dr Paul Yves Sondaar (1934-2003), from the University of Utrecht, the Netherlands, who was the driver behind island research in palaeontology. Paul Sondaar took us with him to many islands for excavations, collection visits and conferences, where he explained us his hypotheses and theories. He coined the term low-gear locomotion for the island ruminants with short and massive limb bones.
The data and results of 40 years of island research, which form the basis for this book, could not have been gathered without the help and assistance of colleagues, collaborators, assistants and curators in the field, the lab, the collections and museums.
The separate chapters were reviewed by the following experts in the field, in alphabetical order, for which we are greatful: the late Larry Agenbroad, Athanassios Athanassiou, Chiara Angelone, Laura Bonfiglio, Pere Bover, the late Johanna de Visser, Lawrence Heaney, Christine Hertler, Mark Lomolino, Ross MacPhee, Sandra Olsen, Hidetoshi Ota, Hiroyuki Otsuka, Maria Rita Palombo, Lorenzo Rook, Haruo Saegusa, Ian Tattersal, George Theodorou, Gert van den Bergh, Lars van den Hoek Ostende, Jan van der Made, and Gerard Willemsen. We thank every one of them for variously commenting on draft and final material, supplying answers to queries and sharing with us their most recent papers.
We are further indebted to the following colleagues (in alphabetical order), who provided us with up-to-date information and relevant papers and with whom we had valuable discussions on various aspects of island mammals throughout the years: Paul Albers, Josep Alcover, Fachroel Aziz, Danilo Balete, Peter Ballmann, Angel Bautista, Rafael Borroto-Paez, Hans Brinkerink, Massimo Delfino, Eusebio Dizon, Clare Flemming, Nuria Garcia, Steven Goodman, Victoria Herridge, Thomas Ingicco, Josephine Joordens, Tassos Kotsakis, Adrian Lister, Raquel Lopez Antonanzas, Federica Marcolini, David Mayhew, Hanneke Meijer, Shai Meiri, Gian Luigi Pillola, Pasquale Raia, David Reese, Jelle Reumer, Roberto Rozzi, Marina Sotnikova, Rab Sukamto, Keiichi Takahashi, Bartholomeus van der Geer, Boris Villier and Rebekka Volmer. Sadly, between the first edition and this revision, Larry Agenbroad, Johanna (Hannie) de Visser, Danilo (Danny) Balete and William (Bill) Stanley passed away. We greatly miss the discussions we had with them on island mammals.
